|
|
|
Fish
Chapter Table Of Contents
Section Objectives
l Describe the distinguishing characteristics of vertebrates.
l Explain how fishes carry out their essential life functions.
l Describe the three basic groups of fishes and give an example of each.
Sites: NetVet Fish --- Ichthyology Links --- Fishes Classes and Orders --- Diagram: Evolution of Fish --- Fish of Lake Michigan --- Fish of Lake Superior --- FishBase: Global Information Database About Fish ---
The name Earth is not particularly appropriate for the planet on which we live, for more than two thirds of its surface is water, not earth. And just about anywhere there is water, there are fishes. At the edge of the ocean, blennies jump from rock to rock and occasionally dunk themselves in tide pools. Beneath the arctic ice live fishes whose bodies contain a biological antifreeze that prevents them from freezing solid in sea water colder than 0°C. And in shallow desert streams in the southwestern United States, pupfish tolerate temperatures that would cook almost any other animal.
To clearly understand what fishes are and how they are related to other vertebrates, it is necessary to know something about the characteristics that unite fishes, amphibians, reptiles, birds, and mammals into the subphylum Vertebrata. You may recall from Chapter 29 that vertebrates belong to the phylum Chordata. This means that fishes and other vertebrates have at some time during their development a notochord, a hollow dorsal nerve cord, and pharyngeal slits. In most vertebrates, the notochord is replaced during development by a backbone, or vertebral column, which encloses and protects much of the nerve cord. In addition, most vertebrates have two sets of paired appendages, a closed circulatory system with a ventral heart, and either gills or lungs for breathing.
1) Chordates are named for their notochord, a semi-flexible rod of cartilage running along the length of the animal. A notochord is present in embryos of all chordates. Some species retain the notochord as adults, but it is replaced by the vertebral column in many species. The spongy material in between the vertebral bones is all that is left of your notochord.
2) Chordates have a hollow dorsal nerve cord which runs above the notochord. Pairs of nerves branch out from the nerve cord, which is enlarnged into a brain in many species. You may refer to your dorsal nerve cord as your “spinal cord.” The dorsal nerve cord and brain make up your central nervous system.
3) Chordates have pharyngeal slits near the pharynx (the area just behind the mouth). They were used for filtering food particles from water in early chordates, later evolving into gills used for breathing. Fishes are examples of chordates that retain their pharyngeal slits (and gills) for life. Amphibians such as frogs have gills when young but lose them as they mature. Some chordates, such as mammals and birds, only have pharyngeal slits during the embryonic stage; yours vanished even before you were born.
4) All chordates have an extension of the notochord and nerve cord that extend past the anus. It’s called a tail.
Some chordates lose their notochords when they become adults. Vertebrates not only keep their notochords—they protect them with bone or cartilage. In most vertebrates, this strucutre is composed of bones called vertebrae. Each bone in your spine is a vertebra. Each vertebra has a hole through which the spinal cord passes. Sharks, skates and rays do not have true bone, but the cartilage in their teeth and vertebrae may calcify, or harden, into a bone-like structure
Fishes can be defined as aquatic vertebrates that are characterized by scales, fins, and pharyngeal gills. However, fishes are so varied that for almost every general statement made about them there are exceptions. For example, some fishes do not have scales. One reason for the many differences among fishes is that four living classes of vertebrates make up the group of animals we know as fishes. Thus many fishes-sharks and lampreys, for example--are no more closely related to one another than humans are to frogs.
There are so many fishes, living and extinct, that their correct scientific classification is complicated. For our purposes, we can say that the living fishes fall into three main groups: jawless fishes, sharks and their relatives, and bony fishes. Sharks and their relatives are also known as cartilaginous fishes because their skeletons are made up of soft, flexible cartilage rather than bone.
Fishes are considered to be the most primitive living vertebrates. (This means that fishes were the first vertebrates to evolve and that they have many characteristics that are thought to have existed in their earliest ancestors. It does not mean that they are somehow inferior to other types of vertebrates.) Fishes did not evolve from such organisms as living lancelets or tunicates. But similarities in structure and embryological development show that fishes and modern invertebrate chordates probably did evolve from common invertebrate ancestors that lived many millions of years ago.
The first fishes--which are also the first vertebrates--were odd-looking jawless creatures whose bodies were covered with bony plates. They lived in the oceans of the late Cambrian Period, about 540 million years ago. For over 100 million years, fishes retained the basic armored jawless body plan. Then during the Ordovician and Silurian periods, fishes underwent a major adaptive radiation. Some of the groups that emerged from this adaptive radiation were jawless fishes that had very little armor--the ancestors of modern lampreys and hagfish. Others were armored jawless fishes in a variety of new forms. These fishes were ultimately evolutionary dead ends that became extinct around the end of the Devonian Period, about 350 million years ago. Still others were armored fishes that possessed a feeding adaptation that would revolutionize vertebrate, evolution: These fishes had jaws.
Jaws are extremely important evolutionary innovations. Jawless fishes are limited to eating small particles of food that they can filter out of the water or suck up like a vacuum cleaner. Jaws made it possible for vertebrates to nibble on plants, munch on other animals, and defend themselves by biting.
Another evolutionary innovation seen in the early jawed fishes were paired pectoral (anterior) and pelvic (posterior) fins that were attached to girdles of cartilage or bone. These fins gave the fishes more control over their movement in the water. In addition, the pectoral fins and girdle provided the raw material from which evolution shaped the forelimbs and shoulder bones of terrestrial vertebrates. Similarly, the pelvic fins and girdle were the origins of the hindlimbs and hip bones.
Although the early jawed fishes soon disappeared, they left behind two major classes that continued to evolve and still survive today. The first of these classes is the cartilaginous fishes, an old and successful group that includes sharks and rays. The second class is the bony fishes, a large and diverse assemblage that contains more than 97 percent of all living fish species.
Form and Function in Fishes
Fishes have entered many environments and evolved adaptations that enable them to survive a tremendous variety of conditions. Here we can give only a brief survey of the many ways fishes accomplish the basic functions of life.
FEEDING Every mode of feeding is seen in fishes--herbivore, carnivore, parasite, filter feeder, and detritus feeder. In fact, a single fish may exhibit several modes of feeding, depending on what kind of food happens to be available. Certain carp, for example, eat just about anything--algae, water plants, worms, mollusks, arthropods, dead fish, and detritus. Some fishes--such as great white sharks, tunas, and barracuda--are carnivores. A few fishes are parasites. For example, pencil catfish live and lay their eggs in the gills of larger fishes. And the male in certain species of anglerfish attaches permanently to the much larger female and obtains nutrients from her blood. Still other fishes, such as lamprey larvae and manta rays, are filter feeders. Although their prey are tiny, many filter feeders are not--the filter-feeding whale shark, which grows as long as 18. 5 meters, is the largest fish in the world.
The adaptations for feeding in fishes are often remarkable. The sawfish (a relative of sharks) kills and stuns prey by slashing into a school of small fish with a long snout edged with sharp teeth. The parrotfish has teeth fused into a short beak that it uses to bite off chunks of living coral and additional teeth in its throat that grind the chunks of coral into sand. The archerfish shoots down insects by spitting drops of water at them. Anglerfish have wormlike or lighted lures that they use to entice prey. And some deep-sea fishes have enormous jaws that allow them to swallow prey larger than themselves!
Although some fishes have strong blunt teeth adapted for crushing clam and other mollusk shells, most fishes do not really chew their food. Instead, they tear their food into conveniently sized chunks or swallow their prey whole
From the mouth, the food passes through a short tube called the esophagus to the stomach, where it is partially broken down. In many fishes, the food is further processed in fingerlike pouches called pyloric ceca (pigh-LOR-ihk SEE-kah; singular: cecum), which are located at the point where the stomach and the intestine meet. The pyloric ceca secrete digestive enzymes and absorb nutrients from the digested food. The intestine receives partially digested food from the stomach and pyloric ceca and completes the process of digestion and nutrient absorption. In the intestine, the digestive enzymes from several other organs, such as the liver and pancreas, are added to the food. Any materials that remain undigested after passing through the intestine are eliminated through the anus.
The structure of a fish's intestine is adapted in ways that help fishes meet their nutritional needs. For example, herbivores typically have a much longer intestine than carnivores. (Incidentally, this is true of most types of animals. ) This gives the animal more time and space to break down plant matter, which is difficult to digest. Lampreys, cartilaginous fishes, and a few bony fishes have a flap of tissue that spirals around the inside of part of the intestine, thus increasing the surface area for nutrient absorption.
RESPIRATION Most fishes breathe with gills that are located on either side of the pharynx. The feathery gill filaments contain many capillaries and provide a large surface area for exchange of oxygen and carbon dioxide. Most fishes breathe by pumping water through the mouth, over the gill filaments, and out through slits in the sides of the pharynx. Some fishes, such as lampreys and sharks, have several gill slits on either side of the pharynx. Other fishes, such as almost all bony fishes, have a single opening through which water is "exhaled. " This opening is usually hidden beneath a protective gill cover.
In many fishes the basic respiratory setup has been modified by evolutionary processes. For example, skates and rays are able to breathe while lying on the ocean floor even though their mouth is located on the underside of their body. Instead of taking in water (and gill-clogging mud and sand) through their mouth, they "inhale" water through special openings on the upper surface of their body.
A number of fishes--such as lungfish, gars, Siamese fighting fish, and kissing gouramis--have a special adaptation that allows them to survive in oxygen-poor water or in areas where bodies of water often dry up. They have specialized organs that serve as lungs by obtaining oxygen from the air. In most airbreathing fishes, this organ is actually a modified swim bladder. A swim bladder, which is found in most bony fish, is a gas-filled sac that lies at the top of the body cavity just beneath the backbone. The majority of fishes use the swim bladder to regulate their buoyancy: Dissolved gases in the blood diffuse into and out of the swim bladder and permit the fish to swim at lesser or greater depths, respectively. Unlike typical fishes, fishes that use the swim bladder as a lung have a tube that connects the swim bladder to the mouth. Some air-breathing fishes are so dependent on getting part of their oxygen from the air that they will suffocate if prevented from reaching the surface of the water.
How in the heck can a fish, which is underwater, breath if there is no air? When we go under water, we have to bring air with us to survive. Whales and dolphins have lungs that store air from the surface. Fish don't have lungs, and they rarely ever venture into the air, so how do they survive. We all know it has something to do with gills, but what exactly.
The water surrounding a fish contains a small percentage of dissolved oxygen. In the surface waters there can be about 5 ml. of oxygen per liter of water. This is much less than the 210 ml. of oxygen per liter of air that we breath, so the fish must use a special system for concentrating the oxygen in the water to meet their physiological needs. Here it comes again, a counter current exchange system, similar to the one we found in the fish's swim bladder and in the tuna's muscles.
The circulation of blood in fish is simple. The heart only has two chambers, in contrast to our heart which has four. This is because the fish heart only pumps blood in one direction. The blood enters the heart through a vein and exits through a vein on its way to the gills. In the gills, the blood picks up oxygen from the surrounding water and leaves the gills in arteries, which go to the body. The oxygen is used in the body and goes back to the heart. A very simple closed-circle circulatory system.
The gills: the gills are composed of a gill arch (which gives the gill rigid support), gill filaments (always paired), and secondary lamellae, (where gas exchange takes place).
Swim Bladders
Bony fish have swim bladders to help them maintain buoyancy in the water. The swim bladder is a sac inside the abdomen that contains gas. This sac may be open or closed to the gut. If you have ever caught a fish and wondered why its eyes are bulging out of its head, it is because the air in the swim bladder has expanded and is pushing against the back of the eye. Oxygen is the largest percentage of gas in the bladder; nitrogen and carbon dioxide also fill in passively.
Physoclistous- swim bladder is closed to the gut. The gas gets in through a special gas gland in the front of the swim bladder. Gas leaves the bladder through an oval body in the back of the swim bladder. The system works in a pretty miraculous way. Oval body, filled by venous blood -gasses leave here
Gas gland, fed by arterial blood -gasses enter here
inside the spots= giant secretory cells- secrete lactate -in capillary clusters rete mirabile
Increased lactate levels from the giant secretory cells lower the surrounding pH, causing the blood hemoglobin to dump off its oxygen. The oxygen diffuses back into the incoming capillary, increasing the partial pressure of oxygen in the incoming capillary. This continues until the partial pressure of the oxygen in the capillary is higher than that of the swim bladder (which has a high concentration of oxygen). This complex system is necessary because the concentration of oxygen is higher in the swim bladder than it is in the blood, so simple diffusion would tend to pull the oxygen out of the bladder instead of pushing it in. If the fish wants more buoyancy, it must tell its secretory cells to release more lactate. Since oxygen diffuses easily with oxygen-poor venous blood, the gas can be forced out.
*Fish that migrate vertically tend to have high oxygen levels in their bladders because it fills in faster and leaves faster.
*Fish that maintain a stable depth tend to have more nitrogen because it is inert, enters slowly, and exits slowly.
INTERNAL TRANSPORT Fishes typically have closed circulatory systems with a heart that pumps blood around the body. The heart consists of two muscular pumping chambers: an atrium (AY-tree-uhm; plural: atria) and a ventricle (VEHNtrihk-uhl). Blood from the body collects in the atrium, which pumps blood into the ventricle. The ventricle pumps blood out of the heart into a muscular vessel called the aorta (ay-OR-tah). Blood goes directly from the aorta into the fine capillary networks in the gills, where gas exchange occurs. From the gills, blood travels throughout the rest of the body tissues and internal organs. As blood leaves muscles and organs, it collects in veins that gather in a thin-walled sac called the sinus venosus (SIGH-nuhs veh-NOH-suhs). From this sinus, blood enters the atrium once again.
EXCRETION Like many other aquatic animals, most fishes get rid of nitrogenous wastes in the form of ammonia. Some wastes diffuse through the gills into the surrounding water. Others are removed by the kidneys, which are excretory organs composed of many tubules that filter nitrogenous wastes from the blood and concentrate them.
Kidneys help fishes control the amount of water in their body. Because fishes in salt water tend to lose water by osmosis, the kidneys of marine fishes concentrate nitrogenous wastes and return as much water as possible to the body. The kidneys of freshwater fishes, on the other hand, pump out lots of dilute urine because in fresh water, a great deal of water continually enters by osmosis. One of the factors that determines which fishes are able to move from fresh to salt water (as salmon do) is their ability to control kidney function.
RESPONSE Fishes have a fairly well-developed nervous system organized around a brain. Fish brains, like those of other vertebrates, have several clearly visible parts.
The most anterior parts of a fish's brain are the olfactory bulbs, which are connected by stalks to the two lobes of the cerebrum (suh-REg-bruhm). In fishes, the cerebrum is primarily involved with the sense of smell, although it also seems to be involved in such behaviors as taking care of young and exploring the environment. The optic lobes process information from the eyes. The cerebellum (ser-uh-BEHL-uhm) coordinates body movements. The medulla (mih-DUL-ah) controls many internal organ functions and maintains balance.
Posterior to the brain is the spinal cord, which is in fact the hollow dorsal nerve cord that characterizes chordates. In cartilaginous and bony fishes, the spinal cord is enclosed and protected by the vertebral column. Between each set of vertebrae, a pair of spinal nerves exits the cord and connects with internal organs and muscles.
Most fishes have superbly designed sense organs that collect information about their environment. Almost all fishes active in daylight have well-developed eyes and color vision at least as good as our own. Fishes active only at night or in cloudy water have large eyes with big pupils that gather as much light as possible. These species do not see color well, but they see in the dark as well as cats do.
Many fishes possess extraordinary senses of taste and smell. Special cells called chemoreceptors (chemo- means chemical) are located all over the head and much of the rest of the body surface as well as in the nose and mouth. Many species, such as catfish, also carry chemoreceptors on their "whiskers. " Salmon can distinguish between the odor of their own home stream and that of another stream while they are still far out at sea. And sharks can detect the presence of a drop of blood in 115 liters of sea water!
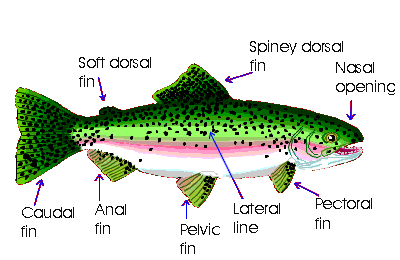
Most fishes have ears inside their head, but they cannot hear sounds well. They can, however, easily detect gentle currents and vibrations in the water. All around their head and down the sides of their body are a series of pores connected to canals beneath the skin that form a sensitive motion detector called the lateral line system. Fishes use their lateral lines to detect other fishes or prey swimming nearby.
In addition to having the senses that we are familiar with, some fishes--electric eels, catfish, and sharks, for example-are able to detect electricity. These electrical detectors are extremely sensitive: A shark can detect one millionth of a voit, which is less than the charge produced by the nerves in an animal's body. Electric eels and electric catfish produce a strong electrical field around their bodies that responds to the presence of nearby objects. Thus electric fishes can locate prey and avoid obstacles in murky water. In addition, electric fishes can produce jolts of electricity (up to 650 volts!) that stun or kill prey and strongly discourage predators.
Successful survival in any environment depends upon an organism's ability to acquire information from its environment through its senses. Fish have many of the same senses that we have, they can see, smell, touch, feel, and taste, and they have developed some senses that we don't have, such as electroreception. Fish can sense light, chemicals, vibrations and electricity.
Light: photoreception [Vision]. Fish have a very keen sense of vision, which helps them to find food, shelter, mates, and avoid predators. Fish vision is on par with our own vision; many can see in color, and some can see in extremely dim light.
Fish eyes are different from our own. Their lenses are perfectly spherical, which enables them to see underwater because it has a higher refractive index to help them focus. They focus by moving the lens in and out instead of stretching it like we do. They cannot dilate or contract their pupils because the lens bulges through the iris. As the depth at which fish are found increases, the resident fish's eye sizes increase in order to gather the dimmer light. This process continues until the end of the photic zone, where eye size drops off as their is no light to see with. Nocturnal fish tend to have larger eyes then diurnal fish. Just look at a squirrelfish, and you will see this to be so. Some fish have a special eye structure known as the Tapetum lucidum, which amplifies the incoming light. It is a layer of guanine crystals which glow at night. Photons which pass the retina get bounced back to be detected again. If the photons are still not absorbed, they are reflected back out of the eye. On a night dive, you may see these reflections as you shine your light around!
Chemicals: chemoreception [Smell and Taste]. Chemoreception is very well developed in the fishes, especially the sharks and eels which rely upon this to detect their prey. Fish have two nostrils on each side of their head, and there is no connection between the nostrils and the throat. The olfactory rosette is the organ that detects the chemicals. The size of the rosette is proportional to the fish's ability to smell. Some fish (such as sharks, rays, eels, and salmon) can detect chemical levels as low as 1 part per billion.
Fish also have the ability to taste. They have taste buds on their lips, tongue, and all over their mouths. Some fish, such as the goatfish or catfish, have barbels, which are whiskers that have taste structures. Goatfish can be seen digging through the sand with their barbels looking for invertebrate worms to eat and can taste them before they even reach their mouths.
Vibrations: mechanoreception [Hearing and touch]. Have you ever seen a fish's ear. Probably not, but they do have them, located within their bodies as well as a lateral line system that actually lets them feel their surroundings.
Fish do not have external ears, but sound vibrations readily transmit from the water through the fish's body to its internal ears. The ears are divided into two sections, an upper section (pars superior) and a lower section (utriculus) The pars superior is divided into three semicircular canals and give the fish its sense of balance. It is fluid-filled with sensory hairs. The sensory hairs detect the rotational acceleration of the fluid. The canals are arranged so that one gives yaw, another pitch, and the last- roll. The utriculus gives the fish its ability to hear. It has two large otoliths which vibrate with the sound and stimulate surrounding hair cells.
Fish posses another sense of mechanoreception that is kind of like a cross between hearing and touch. The organ responsible for this is the neuromast, a cluster of hair cells which have their hairs linked in a glob of jelly known as 'cupala'. All fish posses free neuromasts, which come in contact directly with the water. Most fish have a series of neuromasts not in direct contact with the water. These are arranged linearly and form the fishes lateral lines. A free neuromast gives the fish directional input.
A lateral line receives signals stimulated in a sequence, and gives the fish much more information (feeling the other fish around it for polarized schooling, and short-range prey detection 'the sense of distant touch').
Electricity: electroreception. Sharks and rays posses special organs for detecting electrical potential [voltage]. A set of pits comprise the electroreceptive system called the ampullae of Lorenzini. These are canals in the skin filled with a gelatin-like material that also contain sensory cells. Movements or disturbances near the shark change the voltage drop along the canals, which allows the shark to sense other organisms nearby. These sensors are so sensitive that if there were not any other distortions a shark could detect the heartbeat of a fish 500 miles away! They can detect muscular contractions of struggling prey and even the earth's magnetic field (which sharks use for navigation).
REPRODUCTION In most species of fishes, there are separate males and females. A number of fishes, however, are born as males but change to females as they grow older. Others start as females and later change into males. Unlike many invertebrates, few fishes function as both a male and a female at the same time.
Many fishes are oviparous (oh-VlHP-ah-ruhs), which means they lay eggs. Most oviparous fishes have external fertilization. However, a few oviparous fishes--certain species of sharks and rays, for example--have internal fertilization and thus lay fertilized eggs.
Some oviparous fishes, such as cod, do not take care of their young. Such fish typically release hundreds or even millions of eggs, which increases the chances that a few offspring will survive to adulthood. Other oviparous fishes care for their offspring. For example, Siamese fighting fish build nests of bubbles, and sticklebacks build nests of twigs. Some cichlids and catfish hold their eggs and young in their mouths. And male seahorses hold fertilized eggs in a pouch until the eggs are ready to hatch. These species lay fewer eggs because the parental care means more of the young are likely to survive.
In some species of fishes that have internal fertilization, such as guppies, the eggs develop inside the female. The developing embryos, like those of oviparous fishes, are nourished by food stored in an attached yolk sac. Thus the young do not receive food directly from the mother's body. The young are typically "born" after they have absorbed the yolk sac and are ready to swim on their own. Species with this pattern of reproduction are ovoviviparous (oh-voh-vigh-v!P-ah-ruhs). In other species, including several sharks, the young are actually nourished by the mother's body as they develop. These fishes are said to be viviparous, or truly live-bearing.
Many fishes, including several species you can keep in a home aquarium, exhibit fascinating mating behaviors. Guppy males dance up and down in front of females, trying to get the females interested in them. Cichlids often display beautiful colors to one another and build elaborate nests to attract a mate. A male stickleback will build a nest, drive all other males away, and perform an elaborate dance that shows passing females where his nest is.
| Modes of Reproduction |
|
In fishes, oviparity is most common; the eggs are inexpensive to produce, and as eggs are in the water, they do not dry out (oxygen, nutrients are not scarce). The adult can produce many offspring, which they broadcast into the plankton column. When the offspring settle out of the plankton, they may be in totally new environments, allowing for a great area in which the young may survive. This mode also comes with its disadvantages; when born, the fish must first go through a larval stage for growth before they transform into the adult stage. In this larval stage, they must fend for themselves in obtaining food and avoiding predation. They may not find a suitable environment when they settle out of the plankton column. The survival of individual eggs is very low, so millions of eggs must be produced in order for the parent to successfully produce offspring. The other modes have their advantages, the eggs are much less prone to predation when carried within the mother, and the young are born fully advanced and ready to deal with the environment as miniature adults. These advantages come with a price-tag also; the adult must supply nutrients to its offspring and can only produce a few eggs at a time. The young are limited to the environment that their parents were in, and if this environment is deteriorating, they are stuck with it.
Parental care: In fishes, parental care is very rare as most fish are broadcast spawners, but there are a few instances of parental care. Male gobies guard the eggs in a nest until they are born. The male yellowhead jawfish actually guards the eggs by holding them in his mouth! Weird Fish Sex!
Some fish are very kinky creatures by human standards, displaying behavior that would probably get a human incarcerated for a long time.
FISH SCHOOLING Everyone has heard of a school of fish, an aggregation of fish hanging out together; but why, they are obviously not learning reading, writing, and arithmetic. Schools of fish may be either polarized (with all the fish facing the same direction) or non polarized (all going every which way)
There are some factors that can make it advantageous to hang out with other fish.
Antipredator: by hanging out with other fish, each individual fish may gain an advantage in not being eaten by other fish.
B. Dilution affect. If a fish hangs out with a lot of other fish and a predator does come around, the predator must usually select one prey item. With so many choices, the chances are that it will not be you. This is known as the 'selfish herd'.
C. Predator detection. A bunch of fish has many times the eyes and other senses than a solitary fish; so a school of fish may be better at detecting predators. But a school may also attract predators due to its large size.
Spawning Aggregation: Many fish species form schools only when it comes time to mate. They will form a huge school and release their eggs and sperm in mass quantities. Releasing a massive onslaught of fertilized eggs in the water may be advantages over a solitary egg, because a massive onslaught may be enough to overwhelm the egg predators. The predators will eat as many as they can, but some eggs will inevitably survive.
Enhanced Foraging: A school of fish may have better abilities to acquire food. With many more eyes to detect food, many more meals may be found; but there would also be many more mouths to feed. By working as a team, the school may be able to take larger food items than any one individual could manage to capture.
Migration: The migration abilities of fish in schools may possibly be enhanced due to better navigation, etc. Hydrodynamic efficiency: Due to the complex hydrodynamic properties of water (properties the fish probably discovered only by accident), a fish may gain a swimming advantage by being in a school. The slipstream from the fish ahead of it may make it easier to pass through the water. Good for all the fish except for the ones in front.
Diagram Hagfish vs Lamprey --- Lamprey Dissection Mount
The jawless fishes alive today are divided into two classes: lampreys and hagfishes. Although modern jawless fishes are thought to have evolved from heavily armored, bony ancestors, both lampreys and hagfishes have no bones at all. In fact, they are the only vertebrates that do not have backbones as adults! Instead, their long, snakelike bodies are supported by a notochord.
General Characteristics:

Lampreys are typically filter feeders as larvae and parasites as adults. An adult lamprey's head is taken up almost completely by a circular sucking disk with a round jawless mouth in the center. Adult lampreys live by attaching themselves to fishes (and occasionally whales and porpoises) and scraping away at the skin with their large teeth and a strong, rasping tongue. Lampreys then suck up the tissues and body fluids of their host. Lampreys rarely kill their host, but they leave it in a weakened condition with a large open wound that is easily infected.
The Sea Lamprey belongs in a unique class known as the Agnatha class. This class includes the fish that lack jaws. Yet, it is not a true fish because they do not have certain characteristics such as paired fins and jaws (Moyle 1993). They reach lengths of up to 2.5 feet long, with a recorded record length of 3 feet long (Migdalski 1976). The Sea Lamprey is considered an outer parasite, being that it connects itself to other fish. The main characteristic of the Sea Lamprey is its mouth. The mouth is shaped much like a funnel. Along the inside of the funnel there are many sharp teeth which help it to hold onto the side of the other fish. The Sea Lamprey uses its tongue, which is also covered with sharp teeth, to dig a hole into the fish where it then feeds (Kahl 1997).
The Sea Lamprey is native to the North Atlantic Ocean, but today is found in the Great Lakes after the construction of the Welland Canal in 1829, which is used for the shipping industry. Originally, Niagara Falls was a natural barrier in preventing the Sea Lamprey to migrate into the Great Lakes, until the transformation by the construction of the Welland Canal. (Kahl 1997). By 1921, the Sea Lamprey had migrated to Lake Erie, and by 1936, it had reached Lake Michigan (Migdalski 1976). Currently, the Sea Lamprey can be found most in northern Lake Huron. An estimate claims that northern Lake Huron has as many Sea Lampreys as all the other Great Lakes combined. This is so because of the St. Marys River. This river is how the Sea Lamprey gets to Lake Huron, and since the river has a large volume of water flowing through it, it is easy for the lamprey to get carried into the lake (Kahl 1997).
While under approximately 5 years of age, the Sea Lamprey is found in the rivers and streams that branch off of the Great Lakes. When first born, they drift downstream until they get washed into the muddy water. Here, they hide in the soft muddy ground and feeds on algae and detritus from the surface of the mud (Moyle 1993). After about 5 years, each Sea Lamprey will spend about 12 to 18 months in the Great Lakes. After this time period, it moves upstream where it lives in the gravel constructing nests to bear its young (Kahl 1997).
The Sea Lamprey spends 5 to 6 years, which is most of its life, in the larva stage where it is virtually harmless. It lives in the muddy water eating algae and detritus. Here they are black and eyeless. Here, they are at the most chance of being preyed upon as well. Bigger fish eat the lamprey when at its larva stage, being that it is much like a worm . Then at the age of approximately 5 years, they go into a dramatic transformation during late summer and fall, where they develop their large eyes and their mouth with the sharp teeth. Here, they move upstream into the Great Lakes where they feed on the live fish. They tend to feed on lake trout and whitefish. Apparently they prefer the lake trout because its small soft scales are easily removed. While on the side of a fish, it sucks out the blood and other bodily fluids. Larger fish usually survive the lamprey attack, where the average and smaller fish tend to die. It is not unusual to find a fish with healed lamprey scars, especially in the Great Lake areas. It is estimated that a single lamprey will destroy 40 pounds of fish. When they are at full growth, they are preyed upon mostly the same predators as the other fish, i.e. large birds, and people. Around spring time and after about 12 to 18 months as an adult, they move upstream where they are able to make nests in the gravel. Their nests are crescent-shaped and made out of small stones and gravel. After the nests are completed, the lampreys breed, with females laying anywhere from 30,000 to 100,000 eggs. After this process, the adults die, and the new life cycle starts all over
A new study has claimed that the Sea Lamprey contains spinal axons that can push their way forward allowing the lamprey to repair its spinal cord when it is severed. Scientists are now examining how the lamprey is able to do this, hopefully finding a way to repair spinal injuries to humans and other vertebrates as well.
Hagfishes are probably the most primitive vertebrates alive today. They have pinkish-gray wormlike bodies and four or six short tentacles around the mouth. Hagfishes lack eyes, although they do have light-detecting regions scattered around their body. They feed on dead and dying fish by using a toothed tongue to scrape a hole into the fish's side. Hagfishes have some peculiar traits: They secrete incredible amounts of slime, have six hearts, possess an open circulatory system, and regularly tie themselves into half-knots!
Physical Appearance / Anatomy:

Hagfish, as stated above, are tan-colored deep-sea fish that look slightly like eels. Hagfish are very primitive creatures, with no true bones, no advanced internal organs, and no jaws. The only thing supporting their muscular body is a flexible rod of gristle which functions as a spine. The mouth of the hagfish is nothing more than a slit surrounded by 8 feelers (which hagfish 'smell' with). The most unusual feature of the hagfish is its ability to create slime. Hagfish create so much slime that they frequently need to tie themselves in knots to scrape off the excess. The eyes a hagfish have neither lenses nor lids, and are covered by several layers of flesh (and slime), which inhibits their sight so much that they can only detect light and dark.
Hagfish eat dead or dying fish by boring into the fish with its raspy horned 'tongue,' then literally sucking out their innards. Sometimes, hagfish tie themselves in knots (yes, again) for leverage against their prey.
Hagfish
Misc. Information:
Hagfish have literally no culinary value (except in Japan). Hagfish are fished for their skin, which is tough but soft, and is sold worldwide as 'Eel skin.'
Hagfish eggs are from 0.5 to 2 inches long and are layed in groups of 6-14, which are attached to each other (and the ocean floor) by a sticky white substance (see picture to the right). Information about reproductive habits of the hagfish are unknown, mainly because they are seen (and researched) so little.
Habitat:
The North Atlantic hagfish lives in deep-sea waters off the coast of Northeastern US (southeastern Canada) and western Europe
The class Chondrichthyes (cahn-DRIHK-theez)--which contains sharks, rays, skates, and a few uncommon fishes such as sawfish and chimaeras--is an ancient and successful group. The name of this class (chondros means cartilage; ichthys means fish) refers to the fact that all members have an endoskeleton built entirely out of cartilage. Most of them also have toothlike scales covering the skin. These scales make sharkskin so rough that it is possible to use it as sandpaper.
Most of the 225 living shark species have large
curved tails, torpedo-shaped bodies, and rounded snouts with a
mouth underneath. One of the most noticeable characteristics of
sharks is their enormous number of teeth. A typical shark has
about 3000 teeth arranged in 6 to 20 rows. As teeth in the front
rows are worn out or lost, new teeth are continually replacing
them. A shark goes through about 20,000 teeth in its lifetime!
You have probably heard a lot about shark feeding habits. But contrary to what you may have heard, not all sharks are man-eaters. Some sharks are filter feeders; others have flat teeth adapted for crushing the shells of mollusks and crustaceans. And although there are a number of carnivorous sharks large enough to prey on humans, each year more people are killed by lightning than by sharks.
Unlike sharks, which are adapted for swimming rapidly through the water, rays and skates are adapted for living on the ocean floor. Rays and skates are flattened from top to bottom (you can think of them as squashed sharks), and they swim by flapping their large winglike pectoral fins. Most rays and skates reach a maximum length of about 1 meter, but some, such as manta rays, are up to 7 meters in length.
Sharks are animals that are superbly adapted to
their environment. Almost all are carnivores or scavengers,
although the species that live close to the sea floor feed mostly
on invertebrates. Most possess a keen sense of smell, a large
brain, good eyesight, and highly specialized mouth  and teeth. Their bodies are
usually heavier than water, and they do not have an air filled
swim bladder for buoyancy like most bony fishes. All sharks have
an asymmetric tail fin, with the upper lobe being larger than the
lower one. This feature, together with flattened pectoral fins,
and an oil-filled liver compensates for the lack of a swim
bladder. There are 344 known species of sharks living in all
parts of the oceans, from shallow to deep water and from the
tropics to the polar regions. A few even venture into fresh water
and have been found in rivers and lakes. Contrary to popular
belief, most sharks are harmless to humans. Sharks are classified
into eight orders:
and teeth. Their bodies are
usually heavier than water, and they do not have an air filled
swim bladder for buoyancy like most bony fishes. All sharks have
an asymmetric tail fin, with the upper lobe being larger than the
lower one. This feature, together with flattened pectoral fins,
and an oil-filled liver compensates for the lack of a swim
bladder. There are 344 known species of sharks living in all
parts of the oceans, from shallow to deep water and from the
tropics to the polar regions. A few even venture into fresh water
and have been found in rivers and lakes. Contrary to popular
belief, most sharks are harmless to humans. Sharks are classified
into eight orders:
1. Sawsharks (Pristophoriformes), one family, five sp.Live on the bottom in warm temperate or tropical seas. Easily recognized because of tube, blade like snouts. Bear live young.
2. Dogfish Sharks (Squaliformes), three families, 73 sp.Bottom dwelling deep water sharks, distributed worldwide. Bear live young and eat bony fishes, crustaceans, squid and other sharks. Harmless to humans.
3. Angel Sharks (Squatiniformes), one family, 13 sp.Flattened, bottom dwelling sharks. Found on continental shelves and upper slopes of cold temperate and tropical seas. Have very sharp, awl-like teeth that are used to impale small fish and crustaceans.
4. Bullhead Sharks (Heterodontiformes), one family, 8 sp.Live on rocky reefs where there are plenty of cracks and crevices. Found in Pacific and Indian Ocean. Eat invertebrates.
5. Gilled Sharks (Hexanchiformes), two families, five sp.Deep-water, bottom-dwelling sharks. Worldwide distribution. Only shark with six or seven gill slits. Bear live young and eat bony fish, crustaceans, and other sharks.
6. Mackerel Sharks (Lamniformes), seven families, 16 sp.Small, highly diverse order. Found in tropical to cold temperate or even Arctic waters. Oceanic and coastal. Most very large, eat bony fish, other sharks, squid, and marine mammals. Includes the Mako and Great White and the plankton eating Megamouth and Basking Sharks.
7. Carpet Sharks (Otectolobiformes) seven families, 31 sp.Warm tropical to temperate waters. All members except whale shark live on bottom. Flattened. Most eat small fishes and invertebrates. Whale shark is plankton feeder. Some bear live young and others lay eggs.
8. Ground Sharks (Carcharhiniformes) 8 families, 193 sp.Largest order of sharks. Worldwide distribution, temperate and tropical waters. Most live near coast, although some found in deeper waters. Eat bony fishes, other sharks, squid, and small invertebrates. Includes the dangerous Tiger shark.
The skeleton of elasmobranchs is made of cartilage.
The skeleton may be partially calcified to an extent with calcium phosphates and carbonates, particularly in the vertebral column. This calcified cartilage is not true bone.
The cartilage of a shark's skeleton may be important in future cancer research. Shark cartilage contains an active ingredient that has been known to inhibit tumor growth. In addition, sharks rarely develop cancer. c. A shark's cranium is a single compact cartilaginous block which encloses the brain, olfactory, and auditory capsules. Jaws are loosely attached to the cranium.
Vestigial ribs give no support
A shark's heart is a two-chambered S-shaped tube, small in proportion to body size. Blood flows from the heart to the gills and then to body tissues.
Fast-swimming sharks, such as great whites and makos, have a body temperature that can be quite a bit higher than the surrounding water (up to 8*C or 14.4*F higher). This heat is due to the modified circulatory system associated with the red muscle.
As red muscle functions, it generates heat. Muscle-generated heat warms the blood circulating through the red muscle, which then travels back to the heart through veins. Thus, blood returning to the heart from the muscle is warmer than blood traveling from the heart to the muscle.
Due to the nearness of arteries and veins, heat passes from warmer veins to cooler arteries within the shark's body, rather than dissipating to the cooler environment. This modified circulatory system retains heat in the red muscle.
Sharks have low blood pressure. The walls of the pericardium (the membranous sac that encloses the heart) are rigid, creating a suction within the pericardium to maintain the flow of blood. To circulate blood throughout their bodies, many sharks must swim continuously.
Bony fishes make up the class Osteichthyes (ahs-tee-IHKtheez) (oste- means bone). There are more species in this class than in any of the other vertebrate classes. About 40 percent of all vertebrates are bony fishes. Experts estimate that there are somewhere between 15,000 and 40,000 species alive today.
Almost all bony fishes belong to the enormous group called the ray-finned fishes. This group includes everything from guppies to groupers, salmon, and eels. The name ray-finned refers to the thin bony spines, or rays, that are connected by a thin layer of skin to form the fins. These fins are adapted to a wide variety of functions. Stonefishes, scorpionfishes, and lionfishes have fin rays that are modified into poison spines. Flying fishes, on leaping from the water, can glide with winglike pectoral fins. Mudskippers, which spend a lot of time out of the water, have fins that have evolved into a suction cup that the fish uses in climbing. Triggerfishes have dorsal fins that are usually folded but can be locked in an upright position to help wedge the fish into a hiding place.
 Only seven living species of bony fishes are not
classified as ray-finned fishes. These are the lungfishes and the
coelacanth. These fishes are of interest because they give us an
idea of what the lungs and limbs may have been like in the
ancestors of terrestrial vertebrates.
Only seven living species of bony fishes are not
classified as ray-finned fishes. These are the lungfishes and the
coelacanth. These fishes are of interest because they give us an
idea of what the lungs and limbs may have been like in the
ancestors of terrestrial vertebrates.
The six species of lungfishes alive today are found in Australia, Africa, and South America. The African and South American species live in areas that are flooded during the rainy season but are practically baked during the dry season. When water is available, lungfish use their gills to eliminate carbon dioxide, but they get most of their oxygen by gulping air into a simple sac that functions as a lung. During the dry season, lungfish burrow in the mud and enter a dormant state.
The single species of coelacanth (SEE-lah-kanth) alive today, Latimeria, is the only surviving member of the lobefinned fishes, which were quite common in Devonian times. Unlike ray-finned fishes, which have many bones in the bases of their fins, coelacanths have few bones in their fin bases. Several of these bones are clearly homologous to the limb bones of terrestrial vertebrates. Attached to those bones are a few large rays that form the fins. Ancient lobe-finned fishes seemed to have lived in swampy areas where shallow pools alternated with mud flats and sand bars. Like some of the "walking catfishes" alive today, those lobe-finned species probably used their pelvic and pectoral appendages to move from pool to pool. Unlike its predecessors, the modern coelacanth lives in water about 70 to 400 meters deep in a relatively small area of ocean off the western coast of Africa. However, scientists have observed captive coelacanths "walking" on the bottom of their tank, moving their stubby fins in the same way terrestrial vertebrates move their legs.
Coelacanths were thought to have disappeared with the dinosaurs about 70 million years ago. In 1938, however, fishermen sailing in the ocean off the coast of South Africa caught a strange blue fish that was 1. 5 meters long and had stubby fins and a triple tail. The coelacanth was not extinct after all! Scientists were enormously excited to find living coelacanths because these animals represent a fascinating piece of evolutionary history: Coelacanths are the closest thing we know of to the ancestors of all land vertebrates.

Most fish species have pigmentation. Pigment is mostly contained in cells called chromatophores. Most fishes can contract and expand their chromatophores to change colors. Reflective cells called iridocytes can change color rapidly. Species such as cave-dwelling fishes (family Amblyopsidae) lack pigmentation.
Since different wavelengths of light are absorbed at different depths, fishes may appear a different color underwater than they do at the surface.
Coloration may serve as camouflage. Most species of fishes are countershaded. Countershading is a type of camouflage in which the dorsal (top) side is darker than the ventral (bottom) side. The dorsal side of a countershaded fish blends in with the dark ocean depths or ocean bottom when viewed from above. The ventral side blends in with the lighter surface of the sea when viewed from below. Under ideal circumstances, neither the fish's predators nor the fish's prey can differentiate between the countershaded animal and the environment. The coloration of some fishes helps them blend in with their environment. The northern pike (Esox lucius) has coloration that provides camouflage in weedy areas, making it a more successful predator. Some fishes show disruptive coloration, a camouflage in which the color pattern of the animal contradicts the animal's body shape.
In some species of bony fishes, coloration serves as advertisement to other fishes. Some fishes rely on coloration for species recognition and sexual distinction. The stoplight parrotfish (Sparisoma viride) female and male are completely different colors, although they are similar in shape and size. Some species of fishes become brighter in color during breeding season to attract potential mates. In some species, coloration may trigger territorial behavior. After establishing a territory, the male stickleback's (family Gasterosteidae) belly turns red. He then actively defends his territory only from other fish with red bellies, notably other male sticklebacks. A garibaldi's (Hypsypops rubicundus) bright orange color warns other fishes that the garibaldi will defend its territory.
Coloration may change. In some species, coloration changes with age or sex change. Some species show completely different coloration and markings as they grow from juveniles to adults. Some color change may be rapid and temporary. Alarmed fish, for instance, often change color. Some bottom-dwelling fishes change color almost instantly to match the substrate.
Bioluminescence. Some marine species of bony fishes produce light through a chemical reaction process called bioluminescence. Bioluminescence may be used to attract mates, to deter or confuse predators, or to attract prey. Light production may occur through photophores, luminescent cells on the fish, or by light-producing bacteria that live in or on the fish in a symbiotic relationship.
All fishes have fins. Various bony fish families show various degrees of fin fusion and reduction. Fins help stabilize or propel the fish in the water.
Except in the lungfishes and the coelacanth, fins lack bones. In Actinopterygians, fins are supported by structures called rays. Some bony fishes have soft, flexible fin rays. Other bony fishes have spiny, rigid fin rays at the leading edges of the dorsal, anal, and pelvic fins. Both soft and spiny fin rays are modified scales. The spiny fin rays of some species are associated with venom glands. Fishes in the family Scorpaenidae include the stonefish (Synanceja spp.), the lionfish (Pterois spp.), and the scorpionfish (Scorpaena spp.)-some of the most venomous fishes in the world. Glands in the dorsal, anal, and pelvic spines produce a venom that is intensely painful and is occasionally fatal to humans. There are several records of human deaths due to stonefish stings.
Fishes have two kinds of fins: paired fins (pectoral and pelvic) and median fins (dorsal, caudal, and anal).
The paired pectoral fins are usually responsible for turning. In some fishes, pectoral fins are adapted for other functions. Some bony fishes, such as the hawkfishes (Cirrhitichthys spp.), use their pectoral fins to help them stay at the bottom and on reef areas. Mudskippers (family Periophthalmidae) support themselves on land with their pectoral fins. The pectoral fins of flying fishes (family Exocoetidae) are extremely long, an adaptation that allows flying fish to remain airborne for as long as 20 seconds and glide a distance of 150 meters (492 ft.) or more. Some bottom-dwelling fishes such as threadfins (family Polynemidae) have taste buds and touch receptors on their pectoral fins to locate food.
For some fishes, such as wrasses (family Labridae), pectoral fins are the main source of power for swimming.
Paired pelvic fins add stability and are used for slowing some bony fishes. In the clingfishes (family Gobiesocidae), the pelvic fins have modified into a sucking appendage, which help the fishes hold on to stationary objects on the ocean bottom. The dorsal fin may be a single fin or separated into several fins. In most bony fishes, the dorsal fin is used for sudden direction changes and acts as a "keel" to keep the fish stable in the water. In some fishes, the dorsal fin is adapted for other functions. In the anglerfishes (order Lophiiformes), the dorsal fin is modified into a lure that attracts prey. The dorsal fin of remoras (family Echeneidae) is modified into a sucking disc. Remoras cling to large fishes and mammals with this dorsal disc and are carried along as hitchhikers. A remora's dorsal fin is modified into a sucking disc.
An African knifefish (Gymnarchus niloticus) undulates its dorsal fin to move forward or backward. The caudal fin, or tail, is responsible for propulsion in most bony fishes. Caudal fins come in many shapes: rounded, truncate, forked, and lunate.
Eye size and position vary depending on the particular habitat and behavior of the species. In general, deep-water fishes have large eyes, allowing them to absorb as much light as possible in their dark environments. Shallow-water fishes generally have smaller eyes. The pupils of some species of bony fishes, such as eels, contract and dilate depending on how much light is needed. In most species of bony fishes, however, pupils can't contract or dilate. Flounders and other bony fishes in the order Pleuronectiformes have one eye that migrates across the top of their skull during development to lie adjacent to the eye on the other side. The four-eyed fishes (family Anablepidae) have hourglass-shaped pupils that lie at the waterline as the fish swims at the surface. Four-eyed fishes can focus on images above and below the water simultaneously (Lagier, 1962). The half of the pupil above the waterline focuses on images above the water, while the half of the pupil below the waterline focuses on images under water.
Operculum Most bony fishes have a single pair of gill openings. In most bony fishes, the gill openings are covered by a flexible plate called an operculum that protects the gills. Oxygenated water enters through the fish's mouth and is forced over the gills. It exits behind the operculum. Some species of bony fishes such as eels (family Anguillidae) have a pair of gill holes or pores that aren't covered by an operculum.
The nostrils of most bony fishes have no internal connections with the oral cavity. In some bony fishes such as eels, the nostrils' incurrent and excurrent openings are widely separated.
Mouth shape and size are good indications of a bony fish's feeding habits. Most bony fishes have mouths at the front end of the head. Some bottom-feeding species have mouths on the underside of the snout, angled toward the bottom. Some surface-feeding species have mouths that angle upwards. Butterflyfishes (family Chaetodontidae) have thin snouts and small mouths that are useful in reaching food located in crevices and cracks. Some species of bony fishes, like the goatfishes (family Mullidae), have fleshy barbers that fringe the mouth. These barbers are used to detect food.
Hearing
Bony fishes have an inner ear for equilibrium, detecting acceleration, and hearing. There is no external opening to the ear. A f ish's soft body tissue has about the same acoustic density as water; sound waves travel through soft tissue to the ear. There is great variation in hearing sensitivity, bandwidth, and upper frequency limit among bony fish species. The hearing range of the cod Gadus morhua is about 2 to 500 Hz, with a peak sensitivity near 20 Hz-probably typical for most bony fish species that lack the adaptations described below. Some bony fish species have adaptations for enhanced sound reception at higher frequencies. In some fishes, the swim bladder lies against the ear and acts as an amplifier to enhance sound detection.
In some species, such as goldfish (Carassius auratus), a series of small bones connects the swim bladder to the inner ear. The hearing range of the goldfish is about 5 to 2,000 Hz, with a peak sensitivity near 400 Hz. The adaptive significance of hearing in fishes is not well understood, but most bony fish species probably use hearing to detect prey. In bony fishes, frequency range of sound production does not appear to be correlated with hearing sensitivity. Sound in water travels more than four times the speed of sound in air.
Lateral line
Like the ear, the lateral line senses vibrations. It functions mainly in distance perception and detecting low-frequency vibrations and directional water flow. The lateral line system consists of a series of fluid-filled canals just below the skin of the head and along the sides of a bony fish's body. The canal is open to the surrounding water through tiny pores.
The eyesight in some species of bony fishes may be well developed. Goldfish (Carassius auratus) have excellent visual acuity up to 4.8 m (15 ft.) away.
Bony fishes have a basic vertebrate eye, with various structural adaptations. The water's surface can reflect up to 80% of light that strikes it. Bony fishes have large lenses that help retain a large portion of the light entering the eye. In some species, the eye has a reflecting layer called the tapetum lucidum behind the retina. The tapetum lucidum helps reflect light back through the retina a second time. The mudskipper (family Periophthalmidae) and several other species of bony fishes have accurate eyesight both above and below the surface of the water. Some species have eyes positioned for a wider field of vision below or above their bodies. The South American catfish (family Hypophthalmidae) has eyes directed downward. Many species, including the sand divers (family Dactyloscopidae) have eyes directed toward the surface. Due to its hourglass-shaped pupils, four-eyed fishes (family Anablepidae) can focus on images above and below the water simultaneously. The half of the pupil above the waterline focuses on images above the water, while the half of the pupil below the waterline focuses on images under water.
Bony fishes have taste buds inside their mouths. Some species have taste buds along the head and ventral side of the body. Taste perception hasn't been extensively studied in bony fishes. Some species can detect some sensations, such as salty, sweet, bitter, and acid stimuli. Taste may be responsible for the final acceptance or rejection of prey items.
Olfactory cells in the nasal sac detect tiny amounts of chemicals in solution.
The nasal areas and proficiency of the sense of smell vary among species. In general, the sense of smell is well developed in fishes. Species of freshwater eels (family Anguillidae) may detect chemicals in extremely low dilutions. Eels may detect a substance when only three or four molecules have entered the nasal sac. Studies suggest that species of salmons (family Salmonidae) use smell to guide them to their home streams during the breeding season. Many bony fish species use the sense of smell to detect pheromones, chemical substances released by an animal that influence the behavior of members of the same species. Fishes may release pheromones during the breeding season or when alarmed.
Fishes can become sexually mature at various ages, depending on species. Several factors influence sexual maturity, including age, gender, and size. An spawn immediately after birth. Although female dwarf perch receive sperm soon after they're born, they do not bear young for up to a year. Some bony fishes become sexually mature shortly after birth. The western mosquitofish (Gambusia affinis) becomes sexually mature within a year. . Most bony fishes become sexually mature between one and five years. Most bony fishes are in excess of 8 cm (3 in.) before reproducing. It may take ten years or more for some bony fishes to become sexually mature. The eels (family Anguillidae) become sexually mature at 10 to 14 years of age, and the sturgeons (family Acipenseridae) may take up to 15 years to mature.
In general, species of a small maximum size begin reproducing at an earlier age than those with a large maximum size. Age and associated size are major factors in determination of adulthood.
In most species of bony fishes, sperm and eggs develop in separate male and female individuals. Fertilization can be either internal or external. Males and females may look similar, or they may look very different. Male/female differences may include size, coloration, external reproductive organs, head characteristics, and body shape.
Some bony fishes are hermaphrodites: a single individual produces both sperm and eggs. A fish may be a sequential hermaphrodite or a synchronous hermaphrodite. Sequential hermaphrodites are born one sex and change sex sometime during the course of life.
Like most wrasses (family Labridae), sheephead are born female and have the potential to transform into males later in life. Shown here is an adult male California sheephead (Semicossyphus pulcher).Most wrasses (family Labridae) are sequential hermaphrodites. They are born female, grow into sexually mature females, and have the potential to transform into functional males later in life. In many of the wrasses, sex change correlates with social hierarchy and social behavior. In many wrasse species, social structure includes a large dominant male and many smaller, subordinate females. Removing the male from the group triggers the largest female to begin transforming into a male.
Anemone fishes (family Pomacentridae, subfamily Amphiprioninae) are born male and change into females later in life. Some damselfishes (family Pomacentridae) begin life as males and change into females. In some species, females can revert back to males. Some seabasses (famly Serranidae) change from female to male, and are capable of reverting back to female. Synchronous hermaphrodites have both sperm- and egg-producing organs at the same time. In a few species, self-fertilization is possible.
Some species are unisexual. In unisexual species, there is no fusion of sperm and egg. The sperm is necessary to trigger the egg cell to develop into an adult, but the sperm cell ultimately degenerates and does not take any part in heredity. The resulting young always are females, with no trace of paternal characteristics. Thus, unisexual species are entirely female. They mate with males of related species to produce female offspring. Poecitia formosa is an example of a unisexual species. Always female, P formosa mates with male P mexicana or P latipinna.
Depending on the species, bony fishes may have internal or external fertilization.
Various factors may influence bony fish breeding. Changes in the duration of sunlight, or photoperiod, can stimulate some species of bony fishes to begin reproduction. Temperature change may trigger the maturity of bony fishes in temperate and subpolar areas. Other factors that may affect reproduction are presence of the opposite sex, currents, tides, moon stages, and presence of spawning areas.
Reproduction is generally cyclic in bony fishes. The duration of cycles may be as short as four weeks or as long as many years. Some species spawn continuously throughout the spring and summer. Some bony fishes may spawn many times a year. Many bony fishes reproduce once a year until they die. Other bony fishes may reproduce only once during their lifetime. Pacific salmon (family Salmonidae) reproduce only once during their five-year lifespan, then die soon after.
Diadromous fishes must have access to both marine and freshwater systems to complete their life cycle.
Bony fishes show at least three types of embryonic development: oviparous (egg layers), ovoviviparous (egg retainers), and viviparous (live bearing).
In oviparous development, the female releases eggs. The developing embryo is nourished by a yolk sac. Bony fish eggs generally are spherical. Most are 0.4 to 3.0 mm (0.02-0.1 in.) in diameter. Depending on the species, parents (male and/or female) may scatter, hide, guard, or brood eggs. Some bony fishes lay eggs that drift through the water column. Some bony fish eggs have oil droplets that help them float. Some bottom-dwelling bony fishes produce eggs that sink and remain on the ocean bottom. Some eggs may be sticky or may have tendrils that allow them to attach to plants and other living or nonliving materials in the environment. Some species brood eggs in their mouth or on their skin, fins, or gill areas.
Some fishes that have internal fertilization are ovoviviparous. The female retains fertilized eggs in her body. The developing embryo is nourished by a yolk sac formed prior to fertilization, and there is no nutrient connection between the mother and the developing embryos. One example of an ovoviviparous fish is the seahorse (family Syngnathidae). In seahorses, it's the male that is responsible for incubating fertilized embryos. The female seahorse deposits eggs into a pouch on the male's abdomen. The male releases sperm into the pouch, fertilizing the eggs. The embryos develop within the male's pouch, nourished by their individual yolk sacs. After the embryos have developed, the male gives birth to tiny seahorses.
Viviparous development occurs in some fishes with internal fertilization. Fertilized eggs are retained and develop within the female's ovary or uterus. The developing embryo has a nutrient connection with the mother.
There is great variation in the development stage at which offspring are released. Depending on whether fertilization is internal or external and which type of embryonic development occurs, bony fishes may release eggs or sperm, larvae, or even sexually mature adult fishes. Gestation periods vary among species and between individuals within a species. Since bony fishes are cold-blooded, gestation time may vary within a species. The rate at which the embryo develops depends on the water temperature. The gestation period may be as short as a few days or as long as several months. The number of offspring is inversely related to the chance a single egg has to reach maturity

The skeleton of bony fishes is made of bone and cartilage. The skeleton of a bony fish gives structure, provides protection, assists in leverage, and is a site of red blood cell production. Bony fishes have skeletons that are almost completely calcified. The vertebral column, cranium, jaw, ribs, and intramuscular bones make up the bony fish skeleton.

Tail and trunk muscles. The muscles of the tail and trunk consist of a series of muscle blocks called myotomes. The myotomes usually resemble the letter "W" tipped at a 900' angle. The myotomes are separated by connective tissue called myosepta. The horizontal septum separates the myotomes into dorsal (or epaxial) myotomes and ventral (or hypaxial) myotomes. Jaw muscles usually consist of adductor muscles that close the jaw and abductor muscles that open the jaw.
The fin muscles consist of abductor and adductor muscles that move the fins away from and close to the body, and erector muscles that provide stability and flexibility in the fins.
The nervous system of fishes is poorly developed compared to that of higher vertebrates.
A bony fish's brain is divided into three sections: the forebrain, the midbrain, and the hindbrain. The forebrain is responsible for the bony fish's ability to smell. Bony fishes that have a good sense of smell, such as eels, have an enlarged forebrain. The midbrain is responsible for vision, learning, and motor responses. Blind bony fishes, such as blind cavefishes (family Amblyopsidae), have a reduced midbrain. The hindbrain contains the medulla oblongala and cerebellum, which coordinates movement, muscle tone, and balance. Fast-swimming bony fishes usually have an enlarged hindbrain.
The spinal cord and a matrix of nerves serve the rest of the body.

Bony fishes have a heart with two chambers: the atrium and the ventricle. The venous side of the heart is preceded by an enlarged chamber called the sinus venosus. The arterial side of the heart is followed by a thickened muscular cavity called the bulbus arteriosus. The sinus venosus receives unoxygenated blood from the body. A valve at the end of the sinus venosus opens into the atrium. The atrium has thick, muscular walls. The atrium receives unoxygenated blood and pumps it into the ventricle. The ventricle is the largest and most muscular chamber of the heart. When the ventricle fills with blood it constricts and forces the blood through the bulbus arteriosus. The bulbus arteriosus is a valve or series of valves that control blood flow out of the ventricle and into the ventral aorta. Blood passes through the bulbus arteriosus to the ventral aorta. From the ventral aorta, blood flows to the gill filaments, where it is oxygenated. The spleen, kidney, and bones are sites of red blood cell production.
Oxygenated blood flows from the gill filaments to the organs of the head and body. A complex system of arteries, veins, and capillaries circulates blood through the body.
Some tunas (family Scombridae, tribe Thunnini) maintain a body temperature several degrees higher than that of the surrounding water. This heat is due to the modified circulatory system associated with the red muscle. As red muscle functions, it generates heat. Muscle-generated heat warms the blood circulating through the red muscle, which then travels back to the heart through veins. Thus, blood returning to the heart from the muscle is warmer than blood traveling from the heart to the muscle. Due to the nearness of arteries and veins, heat passes from warmer veins to cooler arteries within the fish's body, rather than dissipating to the cooler environment. This modified circulatory system retains heat in the red muscle. A higher body temperature is an adaptive advantage in that it provides extra power for high-speed swimming. A similar modified circulatory system warms the brain and eye of some species of tunas (family Scombridae, tribe Thunnini) and billfishes (family lstiophoridae).
The esophagus in bony fishes is short and expandable so that large objects can be swallowed. The esophagus walls are layered with muscle. Most species of bony fishes have a stomach. Usually the stomach is a bent muscular tube in a "U" or "V" shape. Gastric glands release substances that break down the food to prepare it for digestion. At the end of the stomach, many bony fishes have blind sacs called pyloric caeca. Although the exact function of the pyloric caeca is unknown, it probably functions both in digestion and absorption of food. The pancreas secretes enzymes into the intestine for digestion. The intestine is where the majority of food absorption takes place. The length of the intestine in bony fishes varies greatly. Herbivorous bony fishes generally have long, coiled intestines, and carnivorous bony fishes have short intestines. The digestive system terminates at the anus.
Water enters the gill chamber through the mouth. A bony fish can remain motionless and breathe by pumping water over its gills by opening and closing its mouth. Blood in the gill filaments absorbs oxygen from the incoming water. Water exits through the gill openings located under the operculum. Lungfish must return to the surface to breathe air. A lungfish swallows air to fill up an air sac or "lung." This lung is surrounded by veins that bring blood to be oxygenated. Its gills alone can't keep a lungfish supplied with enough oxygen to live. Other species such as tarpon (family Elopidae) can gulp air at the surface to supplement their oxygen demand. Some species of bony fishes can absorb considerable amounts of oxygen through their skin.
Many species of bony fishes have a gas-filled bladder called a swim bladder. The swim bladder originally developed as an organ of respiration, as evidenced by the "lung" of the lungfishes. In modern bony fishes that possess a swim bladder, the organ serves principally in maintaining neutral buoyancy. 4. In some fishes the swim bladder has adapted to function as a sound amplifier.

Both marine and freshwater fishes must regulate the movement of water across their body surfaces.
The tissues of marine fishes are less salty than the surrounding water. Water is continually leaving the body of a marine fish through its skin and gills. To keep from becoming dehydrated, a marine fish drinks large amounts of water and produces a small amount of concentrated urine. In addition, its gills are adapted to secrete salt.
The tissues of a freshwater fish are saltier than its surrounding environment. Water is continually entering the body of a freshwater fish through its skin and gills. Freshwater fishes do not drink water, and they produce large amounts of dilute urine.
With representatives in almost every body of water on our planet, fishes are vital parts of many biological systems. For many birds and mammals--including seagulls, raccoons, dolphins, and humans--fishes are important foods. As predators and herbivores, fishes help control the populations of the organisms they eat. Humans have learned to take advantage of this aspect of fish biology. Mosquito fish have been introduced into ponds and lakes in places far from their native home in North America because they consume large amounts of insect larvae. And grass carp and cichlids are used to keep waterways clear of plant growth. Since prehistoric times, humans have caught fishes for food (and for recreation). For at least 4000 years, humans have also raised fishes in artificial ponds. Today we are still trying to find the best ways to cultivate food fishes in underwater farms. And we still keep captive fishes to admire their bright colors and exotic forms.
SECTION REVIEW
1. What are the distinguishing characteristics of vertebrates?
2. Discuss ways in which fishes are adapted for three different modes of feeding.
3. Name and describe three different reproductive strategies in fishes.
4. Describe the three basic groups of fishes. Give an example of a member of each group.
5. Why are lobe-finned fishes and lungfishes important to evolutionary biologists?